.svg)




What cellular senescence actually is at a molecular level
Cellular senescence is a state of permanent growth arrest. When a cell becomes senescent, it stops dividing but doesn't die. Instead, it remains metabolically active and begins secreting a complex mixture of inflammatory cytokines, growth factors, and proteases collectively known as the senescence-associated secretory phenotype (SASP). This isn't a passive shutdown. It's an active, energy-demanding process that fundamentally alters the cell's relationship with its surrounding tissue.
Senescence was originally identified as a response to replicative exhaustion, the point at which telomeres become critically short after repeated cell divisions. But it's now clear that senescence can be triggered by multiple stressors:
- DNA damage from oxidative stress or radiation
- Mitochondrial dysfunction
- Oncogene activation
- Chronic inflammatory signaling
The common thread is persistent cellular stress that activates the DNA damage response, particularly through the ATM and p53 pathways, which halt the cell cycle and initiate the senescent program. In young, healthy tissues, senescent cells are rapidly cleared by the immune system. But with age, both the rate of senescence induction increases and the efficiency of immune clearance declines. The result is an accumulation of senescent cells in tissues throughout the body, where their inflammatory secretions create a toxic microenvironment that impairs tissue function and accelerates the aging of neighboring cells.
Where senescence sits within the hallmarks of aging
The hallmarks of aging, as defined by López-Otín and colleagues, include genomic instability, telomere attrition, epigenetic alterations, loss of proteostasis, disabled macroautophagy, deregulated nutrient sensing, mitochondrial dysfunction, cellular senescence, stem cell exhaustion, altered intercellular communication, chronic inflammation, and dysbiosis. Cellular senescence doesn't exist in isolation within this framework. It's mechanistically linked to at least half of these hallmarks, functioning as both an upstream driver and a downstream consequence.
Genomic instability and telomere attrition
DNA damage is one of the most potent triggers of senescence. When DNA double-strand breaks accumulate or telomeres erode to a critical length, the cell activates a persistent DNA damage response. This response, mediated by kinases like ATM and ATR, phosphorylates histone H2AX at sites of damage, forming nuclear foci that recruit repair machinery. If the damage cannot be repaired, the DNA damage response remains chronically active, locking the cell into senescence.
Telomere attrition is particularly effective at inducing senescence because shortened telomeres are recognized as unrepaired DNA breaks, even though they're a natural consequence of replication. This creates a feedback loop: genomic instability drives senescence, and senescent cells, through their inflammatory secretions, can induce DNA damage in neighboring cells.
Mitochondrial dysfunction
Mitochondrial dysfunction and senescence are bidirectionally linked. Dysfunctional mitochondria produce excessive reactive oxygen species, which damage nuclear and mitochondrial DNA, triggering the DNA damage response that leads to senescence. Conversely, senescent cells exhibit impaired mitochondrial function, including decreased respiratory capacity, reduced membrane potential, and increased ROS production.
This mitochondrial dysfunction in senescent cells isn't just a side effect. It actively sustains the senescent state by maintaining oxidative stress and fueling the energy demands of SASP production. Studies show that mitochondrial-targeted antioxidants can delay the onset of senescence in some contexts, though the human data remains limited.
Chronic inflammation and altered intercellular communication
The SASP is the primary mechanism by which senescent cells drive systemic aging. SASP factors include interleukin-6 (IL-6), interleukin-8 (IL-8), tumor necrosis factor-alpha (TNF-α), matrix metalloproteinases, and growth factors like vascular endothelial growth factor (VEGF). These secreted molecules create a pro-inflammatory microenvironment that disrupts tissue architecture, impairs stem cell function, and induces senescence in neighboring cells through paracrine signaling.
This process, often termed "bystander senescence," allows a small number of senescent cells to have outsized effects on tissue health. The SASP is also a major contributor to inflammaging, the chronic low-grade inflammation that characterizes aging and is associated with nearly every age-related disease, from cardiovascular disease to neurodegeneration.
What drives senescence accumulation with age
Senescent cells accumulate during aging because of an imbalance between the rate at which they're generated and the rate at which they're cleared. Several factors contribute to this imbalance.
Increased senescence induction
Aging tissues experience higher levels of the stressors that trigger senescence:
- Oxidative damage accumulates as mitochondrial function declines and antioxidant defenses weaken.
- DNA damage increases due to both exogenous exposures (UV radiation, environmental toxins) and endogenous processes (replication errors, oxidative lesions).
- Telomeres shorten with each cell division in proliferative tissues.
- Chronic metabolic stress, driven by insulin resistance and nutrient overload, activates inflammatory pathways that can induce senescence.
Each of these processes becomes more prevalent with age, increasing the steady-state production of senescent cells.
Impaired immune clearance
In young organisms, senescent cells are recognized and eliminated by natural killer cells and macrophages. This immune surveillance is highly efficient, preventing senescent cell accumulation even when senescence is induced. But immune function declines with age, a process known as immunosenescence. Natural killer cell activity decreases, macrophage function becomes impaired, and the immune system's ability to distinguish senescent cells from healthy cells diminishes. This decline in immune clearance is compounded by the SASP itself, which can suppress immune function and create an immunosuppressive microenvironment that protects senescent cells from elimination.
Metabolic and hormonal shifts
Metabolic dysfunction accelerates senescence through multiple pathways. Chronic hyperglycemia and insulin resistance increase oxidative stress and activate inflammatory signaling through the NF-κB pathway, both of which promote senescence. Advanced glycation end-products (AGEs), which accumulate with age and are elevated in diabetes, can induce senescence in vascular and renal cells.
Hormonal changes, particularly the decline in growth hormone and IGF-1 with age, may also affect senescence dynamics, though the relationship is complex. While lower IGF-1 is associated with longevity in model organisms, it may also impair the clearance of senescent cells by reducing immune function.
Why individual responses to senescence vary
Not everyone accumulates senescent cells at the same rate, and not everyone experiences the same consequences when they do. This variation is driven by genetics, baseline health, and cumulative exposures.
Genetic determinants of senescence susceptibility
Genetic variants influence both the rate of senescence induction and the efficiency of senescent cell clearance:
- Polymorphisms in DNA repair genes affect how quickly cells accumulate the DNA damage that triggers senescence.
- Variants in telomerase genes (TERT, TERC) determine baseline telomere length and the rate of telomere attrition, directly affecting replicative senescence.
- Genes involved in oxidative stress response, such as those encoding superoxide dismutase and catalase, modulate the oxidative burden that drives senescence.
- Immune-related genetic variants affect the efficiency of senescent cell clearance, with some individuals maintaining more robust natural killer cell function into older age.
Baseline metabolic and inflammatory status
Individuals with better metabolic health, lower baseline inflammation, and higher insulin sensitivity accumulate senescent cells more slowly. This is measurable: people with lower hsCRP, better fasting glucose control, and lower visceral fat have fewer markers of cellular senescence in tissue biopsies. Conversely, chronic metabolic stress from obesity, type 2 diabetes, or metabolic syndrome accelerates senescence across multiple tissues. The inflammatory milieu created by metabolic dysfunction not only induces senescence directly but also impairs immune clearance, creating a vicious cycle.
Cumulative stress and environmental exposures
Lifetime exposure to stressors that induce senescence varies widely. UV radiation, air pollution, smoking, and chronic psychological stress all accelerate senescence in different tissues. Early life adversity and chronic stress increase allostatic load, the cumulative burden of physiological dysregulation, which is associated with accelerated biological aging and increased senescent cell burden. Individuals with lower cumulative exposures and better stress resilience maintain lower senescent cell loads and experience slower functional decline.
What the evidence actually shows about targeting senescence
The concept of targeting senescent cells to slow aging has generated significant interest, but the evidence base is still developing. Most of what we know comes from animal models, with human data limited to early-stage clinical trials.
Senolytic drugs in animal models
In mice, the selective elimination of senescent cells using genetic or pharmacological approaches extends healthspan and, in some cases, lifespan. The combination of dasatinib (a tyrosine kinase inhibitor) and quercetin (a flavonoid) has been shown to reduce senescent cell burden in aged mice, improving physical function, reducing frailty, and extending lifespan. Fisetin, another flavonoid, has demonstrated senolytic activity in preclinical studies, reducing senescent cell markers in multiple tissues. These interventions work by targeting the anti-apoptotic pathways that allow senescent cells to resist programmed cell death, particularly the BCL-2 family of proteins.
Human clinical trial data
Human trials of senolytic drugs are in early stages. Small pilot studies have tested dasatinib plus quercetin in patients with idiopathic pulmonary fibrosis, diabetic kidney disease, and age-related frailty, with some showing reductions in circulating SASP factors and modest improvements in physical function. However, these studies are limited by small sample sizes, short durations, and the lack of long-term safety data. The question of whether reducing senescent cell burden in humans translates to extended healthspan or lifespan remains unanswered. Larger, longer trials are needed to determine efficacy, optimal dosing, and potential risks.
Limitations and unknowns
Senescent cells are not uniformly harmful. In some contexts, particularly wound healing and tissue repair, transient senescence plays a beneficial role by limiting fibrosis and promoting tissue remodeling. The challenge is distinguishing beneficial, acute senescence from the chronic, pathological senescence that accumulates with age.
Current senolytic drugs are not perfectly selective, and off-target effects remain a concern. Additionally, the optimal timing and frequency of senolytic interventions are unknown. Continuous suppression of senescence may have unintended consequences, while intermittent "hit-and-run" dosing may be safer but less effective.
Measuring senescence and its downstream effects
Tracking senescence and its impact on aging requires biomarkers that reflect both the senescent cell burden and the systemic consequences of SASP. While direct measurement of senescent cells requires tissue biopsy, several blood-based markers provide indirect signals.
Inflammatory markers, particularly hsCRP, IL-6, and TNF-α, reflect the systemic inflammatory burden driven in part by SASP. Elevated levels are associated with increased senescent cell accumulation in animal models and correlate with accelerated biological aging in humans. Markers of DNA damage, including circulating cell-free DNA and gamma-H2AX, provide insight into the genomic instability that drives senescence. Telomere length, measured in peripheral blood leukocytes, offers a proxy for replicative history and senescence susceptibility, though it's a noisy biomarker with high individual variability.
Metabolic markers reflect the metabolic dysfunction that both drives and is driven by senescence:
- Fasting insulin, HbA1c, and the triglyceride-glucose index provide a window into the metabolic-inflammatory axis that accelerates aging.
- Advanced lipid markers, including ApoB and Lp(a), reflect cardiovascular risk, which is amplified by senescence-driven inflammation.
Epigenetic clocks, which estimate biological age based on DNA methylation patterns, capture the cumulative effects of aging processes, including senescence. Clocks like GrimAge and DunedinPACE correlate with senescent cell burden and predict mortality risk more accurately than chronological age. While these clocks don't measure senescence directly, they integrate the downstream consequences of senescence and other aging hallmarks into a single metric.
Putting senescence science into practice
Understanding cellular senescence within the broader hallmarks of aging framework clarifies why targeting this single mechanism has the potential to address multiple aging pathways simultaneously. Senescent cells drive inflammation, impair tissue function, and accelerate the aging of neighboring cells through their secretory phenotype. They accumulate because of increased induction and impaired clearance, both of which are modifiable through lifestyle and, potentially, pharmacological interventions.
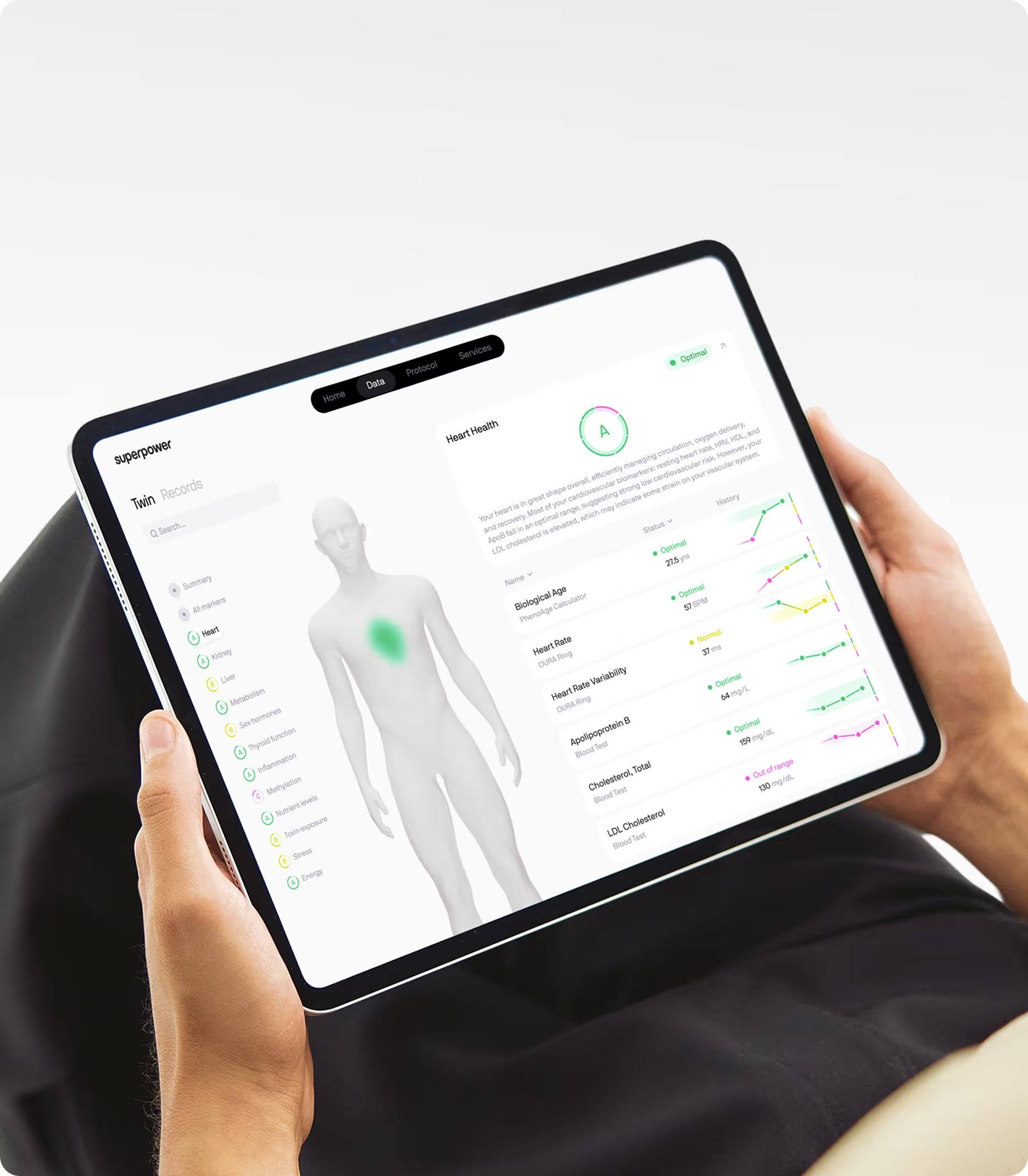
Measuring the markers that reflect senescence burden and its systemic effects provides a baseline for tracking how well interventions are working. Superpower's 100+ biomarker panel includes the inflammatory, metabolic, and cardiovascular markers most relevant to senescence-driven aging, giving you the data to understand where you stand and whether the direction is changing over time.



FAQs
The hallmarks of aging, as defined by López-Otín and colleagues, include genomic instability, telomere attrition, epigenetic alterations, loss of proteostasis, disabled macroautophagy, deregulated nutrient sensing, mitochondrial dysfunction, cellular senescence, stem cell exhaustion, altered intercellular communication, chronic inflammation, and dysbiosis. These processes are mechanistically interconnected, meaning changes in one hallmark can accelerate dysfunction in others, which is why targeting multiple hallmarks simultaneously is a central goal of longevity research.
DNA damage is one of the most potent triggers of senescence. When DNA double-strand breaks accumulate or telomeres erode to a critical length, cells activate the DNA damage response through kinases like ATM and ATR, which halt the cell cycle and initiate the senescent program. Shortened telomeres are recognized as unrepaired DNA breaks, creating a feedback loop: genomic instability drives senescence, and senescent cells, through their inflammatory secretions, can induce DNA damage in neighboring cells.
Senescent cells accumulate because of an imbalance between generation and clearance. Aging tissues experience higher levels of oxidative damage, DNA damage, telomere shortening, and metabolic stress that induce senescence. Simultaneously, natural killer cells and macrophages that normally eliminate senescent cells become less efficient with age due to immunosenescence. The SASP itself can suppress immune function and create an immunosuppressive microenvironment that protects senescent cells from elimination, compounding the imbalance.
The SASP is the complex mixture of pro-inflammatory cytokines, growth factors, and proteases secreted by senescent cells. SASP factors include IL-6, IL-8, TNF-alpha, matrix metalloproteinases, and VEGF. These molecules create a pro-inflammatory microenvironment that disrupts tissue architecture, impairs stem cell function, and induces bystander senescence in neighboring cells. This SASP-driven chronic low-grade inflammation, termed inflammaging, is a major contributor to cardiovascular disease, neurodegeneration, and other age-related conditions.
Dysfunctional mitochondria produce excessive reactive oxygen species that damage nuclear and mitochondrial DNA, triggering the DNA damage response that initiates senescence. Conversely, senescent cells exhibit impaired mitochondrial function, including decreased respiratory capacity and increased ROS production, which actively sustains the senescent state by maintaining oxidative stress and fueling the energy demands of SASP production. This bidirectional relationship means mitochondrial dysfunction and senescence reinforce each other in a damaging cycle.
Senolytic drugs selectively eliminate senescent cells by targeting the anti-apoptotic BCL-2 family proteins that allow senescent cells to resist programmed cell death. In animal models, combinations like dasatinib plus quercetin and fisetin have reduced senescent cell burden, improved physical function, and extended lifespan. Human trials are early-stage: small pilot studies show reductions in circulating SASP factors and modest functional improvements, but long-term safety data and evidence for healthspan or lifespan extension in humans remain limited.
References
- Tartiere, A. G., Freije, J. M. P., & López-Otín, C. (2024). The hallmarks of aging as a conceptual framework for health and longevity research. Frontiers in aging, 5, 1334261. https://doi.org/10.3389/fragi.2024.1334261
- Dodig, S., Čepelak, I., & Pavić, I. (2019). Hallmarks of senescence and aging. Biochemia medica, 29(3), 030501. https://doi.org/10.11613/BM.2019.030501
- Ajoolabady, A., Pratico, D., Bahijri, S., Eldakhakhny, B., Tuomilehto, J., Wu, F., & Ren, J. (2025). Hallmarks and mechanisms of cellular senescence in aging and disease. Cell death discovery, 11(1), 364. https://doi.org/10.1038/s41420-025-02655-x
- Xu, M., Pirtskhalava, T., Farr, J. N., Weigand, B. M., Palmer, A. K., Weivoda, M. M., Inman, C. L., Ogrodnik, M. B., Hachfeld, C. M., Fraser, D. G., Onken, J. L., Johnson, K. O., Verzosa, G. C., Langhi, L. G. P., Weigl, M., Giorgadze, N., LeBrasseur, N. K., Miller, J. D., Jurk, D., … Kirkland, J. L. (2018). Senolytics improve physical function and increase lifespan in old age. Nature medicine, 24(8), 1246-1256. https://doi.org/10.1038/s41591-018-0092-9
- Lopes-Paciencia, S., Saint-Germain, E., Rowell, M. C., Ruiz, A. F., Kalegari, P., & Ferbeyre, G. (2019). The senescence-associated secretory phenotype and its regulation. Cytokine, 117, 15-22. https://doi.org/10.1016/j.cyto.2019.01.013

























.avif)
.svg)
.svg)


