.svg)



Key Takeaways
- Formation mechanism: Polypeptides are formed by ribosomes during translation — the process of reading mRNA and assembling amino acids into a chain, one peptide bond at a time.
- Peptide bond chemistry: Each amino acid is added by a condensation reaction (dehydration synthesis) that releases a water molecule and forms a covalent amide bond.
- Ribosome as catalyst: The ribosome's peptidyl transferase center — an rRNA-based ribozyme — accelerates peptide bond formation approximately ten million-fold over the uncatalyzed rate, enabling typical polypeptide chains to be synthesized in seconds to minutes depending on chain length and cell type — Gerashchenko and colleagues, writing in Nucleic Acids Research in 2021, measured elongation rates of roughly 4–7 amino acids per second across mammalian organs.
- Directionality: Polypeptide chains are assembled from N-terminus to C-terminus, matching the 5' to 3' direction of mRNA reading.
- After synthesis: The newly formed polypeptide must fold, be modified post-translationally, and pass through the cell's quality-control systems before it can function as a protein.
- Laboratory synthesis: Solid-phase peptide synthesis (SPPS) and recombinant expression provide laboratory equivalents of ribosomal synthesis, enabling production of therapeutic polypeptides outside cells.
What Polypeptide Formation Is
Polypeptide formation is the process by which amino acids are joined into a chain. Each connection between adjacent amino acids is a peptide bond — a covalent amide linkage formed by a condensation reaction that releases a water molecule. The sequence of amino acids in the chain — the primary structure — is determined by the genetic information in DNA, transmitted through messenger RNA (mRNA) to the ribosome where assembly occurs. This process is called translation: the nucleotide language of mRNA is translated into the amino acid language of a polypeptide chain. Forbes Kaprive and Krishnamurthy, in the StatPearls peptide biochemistry chapter, define polypeptide formation as a ribosome-catalyzed condensation process that builds the chain from its N-terminus to its C-terminus. Anfinsen, Haber, Sela, and White established the foundational principle that the sequence produced by this process is not random — it is genetically programmed and deterministically encodes the protein's final structure, as demonstrated in their 1961 Proceedings of the National Academy of Sciences paper, showing that sequence determines the three-dimensional structure of the folded protein.
Discovery and history
The molecular mechanism of polypeptide synthesis at the ribosome was clarified progressively through the second half of the 20th century. The Nobel Prize in Chemistry in 2009, awarded to Venkatraman Ramakrishnan, Thomas Steitz, and Ada Yonath, recognized the determination of the atomic-level three-dimensional structure of the ribosome — the machine responsible for all biological polypeptide formation. Nissen and colleagues' 2000 paper in Science established the catalytic RNA basis of the peptidyl transferase center, resolving the long-standing question of whether the ribosome's catalytic activity resided in its protein or RNA components.
How Polypeptides Form in the Body
Polypeptide formation in the body follows a sequence of steps that begins in the cell nucleus with the genetic code and ends at the ribosome with a completed polypeptide chain. The pathway follows the central dogma of molecular biology: DNA is transcribed into mRNA, and mRNA is translated into a polypeptide. Each step in this pathway is actively regulated and subject to multiple fidelity-checking mechanisms.
From DNA to mRNA: the instructions are transcribed
Every polypeptide chain begins with a gene in the cell's DNA. The gene is a specific sequence of nucleotides encoding the amino acid sequence of the polypeptide. During transcription, the enzyme RNA polymerase reads the gene's DNA template strand and synthesizes a complementary mRNA strand. The mRNA is a single-stranded copy of the gene's information, written in the RNA nucleotide alphabet (using uracil instead of thymine). The mRNA is processed — capping, polyadenylation, and splicing of introns — and exported from the nucleus to the cytoplasm where translation occurs. The mRNA sequence is read in triplets: each three-nucleotide sequence (a codon) specifies one amino acid. Sanvictores and Farci establish in the StatPearls primary structure review that the primary amino acid sequence is encoded directly in DNA — a change in one codon at the DNA level produces a different amino acid at that position in the chain. mRNA modifications can also regulate translation: Franco and Koutmou, writing in Biophysical Chemistry in 2022, showed that chemical modifications to mRNA nucleobases affect translation elongation rate, demonstrating that the mRNA is not simply an inert message but an actively regulated template.
Ribosomal translation: assembling the chain
At the ribosome, the mRNA template is read codon by codon and the corresponding amino acids are assembled into the polypeptide chain. The ribosome has three key sites: the A site (aminoacyl), where each new incoming aminoacyl-tRNA brings the next amino acid; the P site (peptidyl), where the growing polypeptide chain is held; and the E site (exit), where the now-empty tRNA departs. Voorhees and Ramakrishnan, writing in the Annual Review of Biochemistry in 2013, described the structural basis of the translational elongation cycle in which the ribosome extends the polypeptide chain codon by codon.
Translation proceeds through three phases. In initiation, the ribosome assembles on the mRNA at the start codon (AUG, encoding methionine) — in eukaryotes every polypeptide therefore begins with methionine at its N-terminus (bacteria use formyl-methionine), though this residue is often removed post-translationally. In elongation, each codon is decoded by an aminoacyl-tRNA bearing the complementary anticodon, the ribosome catalyzes formation of the peptide bond, and the ribosome translocates along the mRNA by one codon. Frank and colleagues, writing in PNAS in 2007, described the mechanochemistry of mRNA-tRNA translocation at the ribosome. In termination, a stop codon (UAA, UAG, or UGA) is recognized by a release factor rather than a tRNA, triggering hydrolysis of the chain from the P-site tRNA and release of the completed polypeptide.
The chemistry of peptide bond formation: dehydration synthesis
The chemistry at the core of polypeptide formation is a condensation reaction — dehydration synthesis — in which a water molecule is released for each peptide bond formed. Specifically, the alpha-amino group of the incoming A-site amino acid attacks the ester bond linking the P-site peptidyl chain to its tRNA. This nucleophilic attack transfers the growing chain from the P-site tRNA to the A-site aminoacyl-tRNA, extending the chain by one residue and releasing a water molecule. Rodnina and colleagues, in their 2006 Quarterly Reviews of Biophysics review, showed that the ribosome accelerates this reaction approximately ten million-fold over the uncatalyzed rate — without this catalysis, assembling even a small polypeptide would require years. Nissen and colleagues established in 2000 that this catalysis is provided by the rRNA component of the large ribosomal subunit, making the ribosome one of the most important ribozymes in biology. Rodnina, Beringer, and Wintermeyer, in a 2007 Trends in Biochemical Sciences review, characterized the ribosome's catalysis as primarily entropic — it lowers the activation entropy rather than the activation enthalpy of the reaction. Hiller and colleagues, writing in Nature in 2011, described the two-step chemical mechanism of ribosome-catalyzed peptide bond formation.
After synthesis: folding, modification, and quality control
A completed polypeptide chain emerging from the ribosome is linear and unfolded. It must reach a specific three-dimensional conformation before it can function as a protein. In the cell, this folding begins co-translationally — while the chain is still being synthesized — and continues after the chain is released from the ribosome. Molecular chaperones in the endoplasmic reticulum bind to exposed hydrophobic regions and prevent aggregation while the chain completes its fold. Adams and colleagues, writing in Progress in Molecular and Subcellular Biology in 2021, reviewed chaperone-assisted co- and post-translational polypeptide folding in comprehensive detail. Ellgaard and colleagues (Braakman group), writing in Traffic in 2016, described the co-translational and post-translational folding environments that shepherd polypeptides toward their native protein conformations. Halperin, Jung, and Michalak, in a 2014 review in IUBMB Life, catalogued the many functions of ER chaperones and folding enzymes — including disulfide isomerases that catalyze correct disulfide bond formation and glycosylation machinery that adds carbohydrate modifications required for certain proteins' final structure and secretion. When folding fails and the quality-control system is overwhelmed, misfolded polypeptides can aggregate — as Stefani and Dobson documented for amyloid-forming polypeptides in Alzheimer's and prion diseases.
Regulation of Polypeptide Formation
Polypeptide formation is not a simple mechanical process — it is actively regulated at multiple levels, from the rate of elongation to the fidelity of codon reading.
Translation rate and elongation pausing
The ribosome does not move along mRNA at a constant rate. Certain codons are decoded more slowly than others, depending on the availability of the corresponding charged tRNA species. Varenne and colleagues, writing in the Journal of Molecular Biology in 1984, documented that tRNA availability affects the rate of elongation of nascent polypeptide chains — a finding with implications for codon optimization in recombinant protein production. Richter and colleagues, writing in Cell in 2015, showed that ribosome pausing during elongation serves regulatory functions including co-translational folding and targeting. Chu and colleagues, writing in The EMBO Journal in 2014, demonstrated that translation elongation rate controls translation initiation on eukaryotic mRNAs — linking the rate of polypeptide formation to the overall level of protein synthesis from a given mRNA.
Ribosomal proofreading and fidelity
Kramer and Farabaugh, reviewing the literature in RNA in 2007, estimated ribosomal missense error rates at roughly one mistake per 1,000 to one per 10,000 codons — a fidelity level that requires active proofreading mechanisms. The small ribosomal subunit monitors the geometry of codon-anticodon base pairing and can reject incorrectly paired tRNAs before peptide bond formation. Zaher and Green, writing in Nature in 2009, characterized ribosomal quality control following peptide bond formation, documenting mechanisms that allow the ribosome to abort synthesis of aberrant chains. This proofreading is the cellular basis for the high structural fidelity of the polypeptides produced from a given genetic template.
Laboratory Polypeptide Synthesis: The In Vitro Alternative
Ribosomal translation is not the only pathway to polypeptide formation. Laboratory synthesis methods produce therapeutic polypeptides outside cells, with precise control over sequence and chemical modifications.
Solid-phase peptide synthesis (SPPS)
SPPS builds polypeptide chains stepwise from C-terminus to N-terminus on a solid resin support, adding one protected amino acid building block per cycle. The method, developed by Bruce Merrifield (Nobel Prize, 1984), allows synthesis of any defined sequence up to approximately 50 amino acids with high purity. Sharma and colleagues, writing in Drug Discovery Today in 2023, described SPPS as the primary method for producing therapeutic peptides that are not accessible through recombinant expression.
Ring-opening polymerization of N-carboxyanhydrides
For longer synthetic polypeptides used in drug delivery and biomaterials research, ring-opening polymerization (ROP) of alpha-amino acid N-carboxyanhydrides (NCAs) provides an abiotic route to high-molecular-weight polypeptide chains. Feng and colleagues, writing in the International Journal of Molecular Sciences in 2022, reviewed synthetic polypeptides produced by NCA ring-opening polymerization and their applications in anti-tumor drug delivery. Zhang and colleagues, writing in Chemical Communications in 2021, described stimuli-responsive synthetic polypeptides produced by this chemistry for controlled drug release applications.
Recombinant expression
For longer polypeptides and proteins that cannot be efficiently assembled by SPPS, recombinant DNA expression in microbial hosts (bacteria, yeast, mammalian cells) allows the cell's ribosomal translation machinery to produce the target polypeptide from a gene construct. This approach is used to produce insulin, erythropoietin, monoclonal antibodies, and other protein biologics. As of April 2026, recombinant expression is the production method for virtually all FDA-approved protein biologics and many polypeptide therapeutics. The Sharma 2023 review cited earlier (PMID 36481586) documents both SPPS and recombinant expression in the landscape of current therapeutic polypeptide production methods.
Why This Matters for Your Health
Understanding polypeptide formation connects the genetic code to clinical outcomes in a direct chain. The accuracy of ribosomal translation maintains the integrity of every protein in your body — the enzymes that regulate your metabolism, the hormones that coordinate your endocrine system, the structural proteins that give your tissues their mechanical properties. When translation accuracy is compromised — by ribosome inhibitors (some antibiotics work this way), by mRNA damage, or by accumulation of misfolded aggregates — the consequences include disease. The misfolded polypeptide aggregates observed in Alzheimer's disease and type 2 diabetes involve the same underlying polypeptide substrates that, when correctly folded, serve normal biological functions — a reminder that disease-associated protein aggregation involves the same molecular materials as healthy physiology.
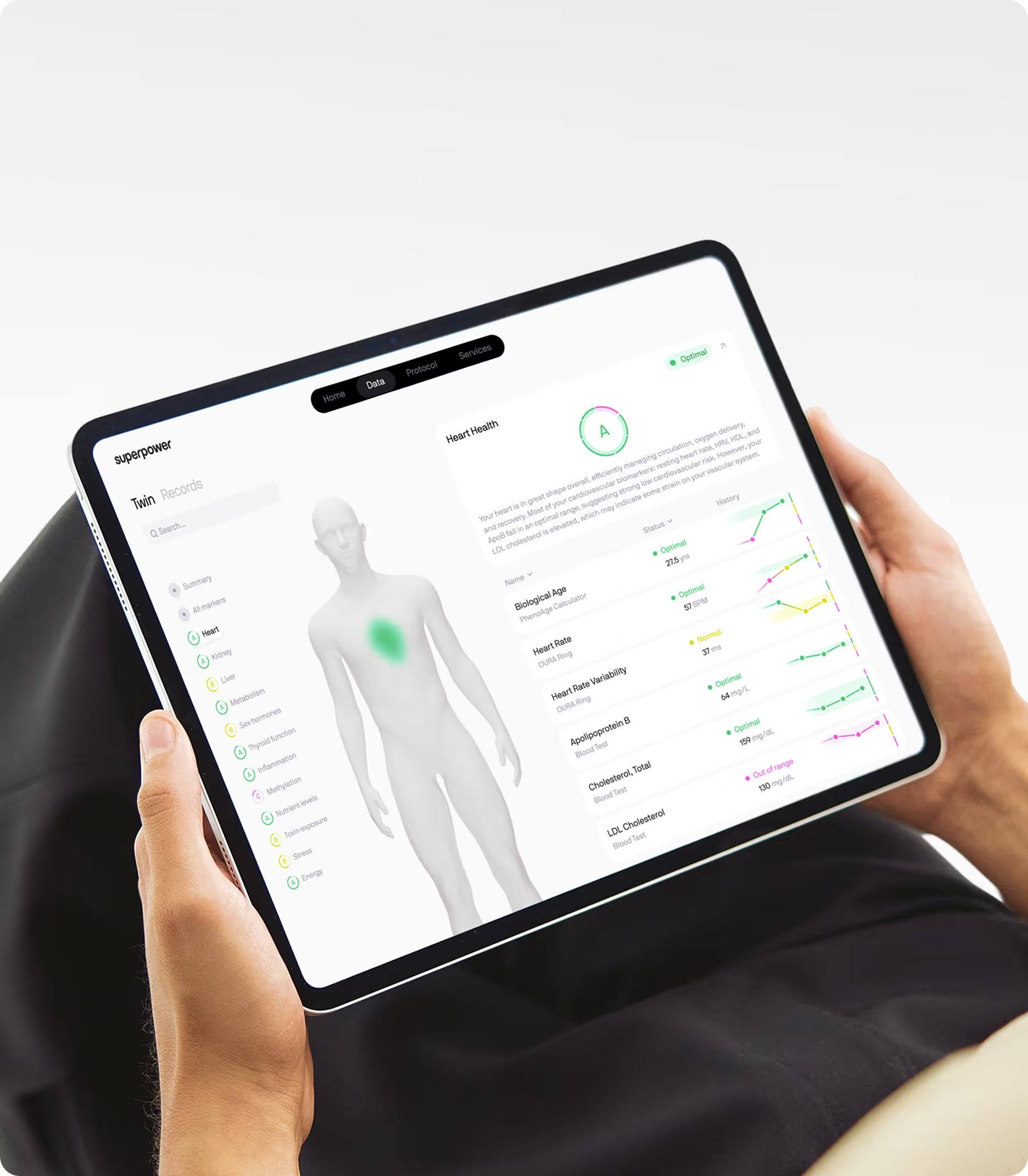
The polypeptide hormones produced by this machinery — insulin, IGF-1, glucagon, and others — are directly measurable in clinical bloodwork. Their levels reflect the fidelity and output of the polypeptide formation process in the cells that produce them. That is the principle at the center of Superpower's preventive health approach: measuring the molecular products of biological processes — including the polypeptide hormones reported on a blood panel — is what gives preventive data its interpretive power.
Which Biomarkers Are Relevant if You Are Exploring Polypeptide Formation?
While the act of polypeptide formation itself is not directly measured, the products of correct and incorrect polypeptide formation are measurable in standard clinical panels. These markers reflect the output of the translation and folding machinery described in this article.
- Insulin: The functional form of insulin — the polypeptide produced by beta-cell ribosomes, processed through the ER, and secreted — is what a fasting insulin blood test measures. Elevated fasting insulin is an early detectable marker of insulin resistance and reflects the metabolic demand on beta-cell polypeptide synthesis.
- IGF-1: A 70-amino acid polypeptide synthesized by liver cells in response to GH signaling. IGF-1 levels reflect both the hepatic production capacity and GH axis activity. It is the standard clinical marker for assessing anabolic signaling and is included in comprehensive metabolic panels.
- HbA1c: A post-translational modification of the hemoglobin polypeptide chain by glucose — a chemical reaction that accumulates over the erythrocyte's lifespan. HbA1c reflects average blood glucose over approximately three months and is the primary long-term glycemic marker in clinical practice.
- hs-CRP: C-reactive protein is a polypeptide synthesized by hepatocytes in response to IL-6 inflammatory signaling. Elevated hs-CRP levels indicate systemic inflammation and correlate with cardiovascular and metabolic risk. It reflects the inflammatory activation of the liver's polypeptide synthesis machinery.
- Total protein: Serum total protein reflects the integrated output of polypeptide synthesis across the body — primarily albumin and globulins. Low total protein may indicate impaired ribosomal production, increased degradation, or inadequate amino acid supply for synthesis.
This page is provided for educational and informational purposes only and does not constitute medical advice. The biological information presented reflects the current scientific literature. Always consult a qualified healthcare provider before making changes to your health routine.



FAQs
In biology, polypeptides are formed inside cells through a process called translation, in which ribosomes read messenger RNA (mRNA) and assemble amino acids into a growing chain, joining each new amino acid to the previous one by a peptide bond. The basic biochemistry is described by Forbes Kaprive and Krishnamurthy in the StatPearls peptide chapter, defining polypeptide formation as a condensation reaction that releases a water molecule at each step. The chain grows from its N-terminus to its C-terminus until a stop codon on the mRNA signals the ribosome to release the completed chain.
A condensation reaction — also called dehydration synthesis — is the chemical reaction that forms each peptide bond during polypeptide assembly. The carboxyl group (C=O–OH) of the incoming amino acid reacts with the amino group (NH₂) of the preceding amino acid, releasing a molecule of water and forming a covalent amide (peptide) bond. Rodnina and colleagues, in their 2006 Quarterly Reviews of Biophysics review, described the chemistry of ribosomal peptide bond formation as a nucleophilic attack by the alpha-amino group of the A-site aminoacyl-tRNA on the ester bond of the P-site peptidyl-tRNA. The ribosome's role is to position the reacting groups and catalyze the reaction — not to supply the chemical energy, which comes from hydrolysis of the aminoacyl-tRNA ester bond.
The ribosome is the molecular machine that reads mRNA and catalyzes each peptide bond formation step. It has two subunits: the small subunit reads the mRNA codon sequence; the large subunit contains the peptidyl transferase center that catalyzes the condensation reaction forming each peptide bond. Nissen and colleagues established in their 2000 Science paper that the catalytic activity of the peptidyl transferase center is provided by rRNA — not ribosomal protein — making the ribosome a ribozyme. Rodnina and colleagues showed that this RNA-based catalysis accelerates peptide bond formation approximately ten million-fold over the uncatalyzed rate.
Dehydration synthesis is the general term for any reaction that forms a covalent bond between two molecules by releasing a water molecule. In polypeptide formation, dehydration synthesis is the specific mechanism by which each peptide bond is created: a water molecule (H₂O) is released for every amino acid added to the growing chain. Because polypeptides are assembled one residue at a time by this reaction, a polypeptide of 100 amino acids requires 99 condensation reactions and releases 99 water molecules during its synthesis.
The N-terminus (amino terminus) is the end of the polypeptide chain that begins with a free amino group (–NH₂) — the first amino acid incorporated during synthesis. The C-terminus (carboxyl terminus) is the end with a free carboxyl group (–COOH) — the last amino acid added. Polypeptide chains are assembled directionally from N-terminus to C-terminus, matching the 5' to 3' reading direction of the mRNA. Sanvictores and Farci, in the StatPearls primary protein structure chapter, describe the N-to-C directionality of polypeptide chain assembly. Protein sequences are always written and reported in this N-to-C direction.






























.avif)
.svg)
.svg)
